

OPERÁRIAS ÓRFÃS:
MUDANÇAS NO COMPORTAMENTO E LONGEVIDADE DA ABELHA CANUDO (Scaptotrigona aff. postica)
Bárbara dos Santos Conceição Lopes*1 – Alistair John Campbell1,2 – Felipe Andrés León Contrera1
1 Instituto de Ciências Biológicas, Laboratório de Biologia e Ecologia de Abelhas, Universidade Federal do Pará, Belém, Pará, Brasil
2 Laboratório de Botânica, Embrapa Amazônia Oriental, Belém, Pará, Brasil
*Correspondência: B. S. C. Lopes – barbaralopes_@hotmail.com
Introdução
A divisão reprodutiva de trabalho em insetos sociais é apresentada em castas, em que a rainha (indivíduo reprodutivo) é sustentada pelos indivíduos não reprodutivos, as operárias (Crespi, 1994). Em abelhas sem ferrão (Tribo Meliponini) a rainha mantém o controle sobre as operárias, principalmente, através da sua presença, mostrando comportamento de dominância, ao invés de controle feromonal (Kleinert, 2005; Nunes et al., 2017; Veiga et al., 2017). Em colônias com rainha, normalmente, as operárias são estéreis, botando apenas ovos tróficos que servem de alimentação para rainha. Entretanto, em algumas espécies de Meliponini, elas podem desenvolver seus ovários e produzir ovos funcionais haploides que originam machos (Sakagami, 1982; Tóth et al., 2002) através de partenogênese.

Nesse sentido, a morte da rainha fisogástrica (poedeira) pode implicar em desorganização no comportamento das operárias. Existem algumas possibilidades de uma colônia perder sua rainha, por exemplo, o desaparecimento dela pode ocorrer por um manejo humano inadequado, que a feriu e provocou a sua morte. Ou ela já era velha e, por razões diversas, não conseguiu criar uma nova rainha virgem para substituí-la.
São poucos os trabalhos que abordam os efeitos da orfandade sobre colônias de abelhas nativas sem ferrão. Em colônias órfãs de Plebeia saiqui e P. catamarcensis as operárias realizam alguns comportamentos atípicos, como construção de células de cria irregulares e oviposição (Imperatriz-Fonseca e Oliveira, 1976; Pinho et al., 2010). Para abelhas operárias, a longevidade está relacionada ao esforço fisiológico desempenhado durante sua vida, considerando o esforço da procura e coleta de recursos, ou mesmo os riscos da predação fora do ninho (Page Jr. e Peng, 2001). As condições internas da colônia também afetam a longevidade, como por exemplo, a presença ou ausência de rainha fisogástrica (Alves et al., 2009).
Nesse contexto, objetivamos comparar o comportamento e longevidade de Scaptotrigona aff. postica em colônias sem rainha (órfãs) e colônias com uma rainha fisogástrica presente.
O estudo foi realizado no meliponário da Embrapa Amazônia Oriental pela bióloga Bárbara dos Santos Conceição Lopes durante o seu mestrado em zoologia na Universidade Federal do Pará/Museu Paraense Emílio Goeldi. A dissertação gerou um artigo feito em parceria com o Dr. Alistair John Campbell e o Dr. Felipe Andrés Léon Contrera, publicada na revista Behavioral Ecology and Sociobiology (em inglês). Link: https://rdcu.be/b2zci.
Quais questões foram abordadas?
Nº1. Existem diferenças na atividade de forrageamento entre os dois tipos de colônias?
Nº2. Existe alguma diferença na idade das operárias que constroem células de cria?
Nº3. A idade de forrageio das operárias é afetada em colônias sem rainha?
Nº4. Aumentando, assim, a expectativa de vida das operárias órfãs?
Como fizemos?

A espécie estudada foi a Scaptotrigona aff. postica, um dos meliponíneos comumente utilizados na meliponicultura (criação racional de abelhas nativas sem ferrão) no Pará, para produção de mel e é cada vez mais usada na polinização de culturas regionais (Leão et al., 2016). Foram escolhidas oito colônias desta espécie e transferidas para caixas de observação (30 cm x 30 cm x 22,5 cm), que foram colocadas em cima de suportes individuais (cavaletes) distanciadas cerca de três metros uma das outras. Todas as colônias foram similares, considerando número de potes de alimento e discos de cria, e tamanho populacional estimado (Figura 1). Dessas oito colônias, quatro continuaram com rainha (CR) e as outras quatro tiveram a rainha fisogástrica e rainhas virgens removidas (sem rainha – SR), além da destruição das células reais (realeira), para evitar o nascimento de novas rainhas virgens. As operárias emergentes (recém-nascidas) foram marcadas com tinta atóxica na região central-dorsal do tórax com cores diferentes por dia de nascimento, formando coortes etárias (grupos de idade) em todas as colônias (CR e SR; Figura 2).
Para responder à pergunta 1, avaliamos a atividade de forrageamento ao longo do tempo (medido em dias) em colônias que perdem sua rainha, comparadas às colônias com rainha presente. Para responder à pergunta 2, relacionamos a idade das operárias órfãs marcadas que participaram da construção de células de cria ao longo do tempo (medido em dia), comparando com as operárias marcadas das colônias com rainha. Para responder à pergunta 3, relacionamos a idade de forrageio das operárias de colônias sem rainha ao longo do tempo (medido em dia), comparadas às colônias com rainha. E para responder à pergunta 4, comparamos as curvas de sobrevivência das colônias sem rainha e com rainha.

Quais os resultados encontrados?
1 – Atividade de forrageamento
As colônias dos tratamentos (CR e SR) mostraram atividade de forrageamento (número de forrageiras) diferentes ao longo do tempo, com as colônias com rainha (CR) apresentando um ligeiro decréscimo na atividade externa. Entretanto, colônias sem rainha tiveram maior redução na atividade de forrageio (Figura 3a). Em média, em colônias SR a atividade de forrageamento foi cerca da metade da encontrada em colônias CR (Figura 3b; Tabela 1).
2 – Idade das operárias envolvidas na construção de células de cria
Houve diferença significativa na idade das operárias envolvidas na construção de células de cria entre as colônias com rainha (CR) e sem rainha (SR). Nas colônias CR as operárias que participaram da construção de células de cria foram apenas as juvenis, já em colônias órfãs observamos que, além das operárias juvenis, as operárias mais velhas também construíram células de cria ao longo do tempo (Figura 3c). Em média as operárias marcadas envolvidas na construção de células de cria em colônias SR apresentaram sete dias de idade a mais do que operárias marcadas que realizaram essa mesma atividade em colônias CR (Figura 3d; Tabela 1).
Quais as consequências para as colônias órfãs?
Nosso estudo mostrou que em S. aff. postica o número de forrageiras em colônias órfãs diminui muito mais do que em colônias com rainha, provavelmente porque em colônias sem rainha a produção de novas operárias (através da oviposição pela rainha), que naturalmente substituem as forrageiras que morrem, tinha cessado (Skagami, 1982). É possível que a condição de orfandade altere o tamanho da população colonial. Nós não mensuramos taxa de postura e de nascimentos para verificar a quantidade de crias ao longo do tempo, mas foi visível em nosso estudo que nas colônias sem rainha o número de abelhas emergindo das células de cria foi diminuindo ao longo do tempo. Acreditamos que isso, provavelmente, deve ter afetado a atividade de forrageio. Em colônias de P. remota, por exemplo, foi observada a atividade de forrageamento diário na fase reprodutiva e diapausa reprodutiva (período sem processo de aprovisionamento de células e oviposição), em que os resultados revelaram que na fase reprodutiva o forrageio é quase constante ao longo do dia, já na diapausa reprodutiva o número de abelhas em forrageio é menor (Nunes-Silva et al., 2010).

Também observamos que nas colônias sem rainha a idade das operárias de S. aff. postica envolvidas na construção de células de cria aumentou ao longo do tempo. Provavelmente, essas operárias órfãs permanecem construindo células de cria na tentativa de produzir seus próprios filhos (machos oriundos de ovos não fecundados). Operárias do gênero Scaptotrigona em colônias com rainha fisogástrica foram observadas fazendo comportamento de postura de ovos, revelando a participação delas na produção de machos, em que 13% a 42% deles são produzidos pelas operárias (Bego, 1982; Paxton et al., 2003; Tóth et al., 2004). Provavelmente, esse comportamento de oviposição pelas operárias órfãs de S. aff. postica intensifique a produção de machos, possibilitando que a colônia sobreviva até o aparecimento de uma nova rainha virgem ou recém-acasalada vinda de outros ninhos, que poderá se estabelecer nessa colônia. Em nosso experimento, próximo da fase final, ainda foi possível visualizar em ninhos sem rainha alguns ovos das operárias em uma célula de cria isolada, contendo até três ovos (Figura 5).
Nossos dados mostram que a perda da rainha também afeta a idade de forrageio, em que algumas operárias de S. aff. postica coletaram recursos em idades mais avançadas e outras de mesma faixa etária (50-70 dias de idade) ficaram dentro do ninho. Em colônias sem rainha de Apis mellifera cerana foi observado que após algumas operárias ativarem seus ovários e começarem a ovipositar, algumas delas passaram a evitar o forrageio ou mudaram para o forrageamento tardiamente (Tan et al., 2015). Acreditamos que, provavelmente, algumas operárias órfãs de S. aff. postica também evitam o trabalho arriscado fora da colônia e permanecem dentro do ninho competindo pela oviposição, como mencionado acima sobre a figura 5. Entretanto, em ninhos órfãos capturamos algumas forrageiras com idade avançada retornando da atividade externa, seja porque estavam forrageando eventualmente ou porque teriam iniciado esse trabalho mais tardiamente.
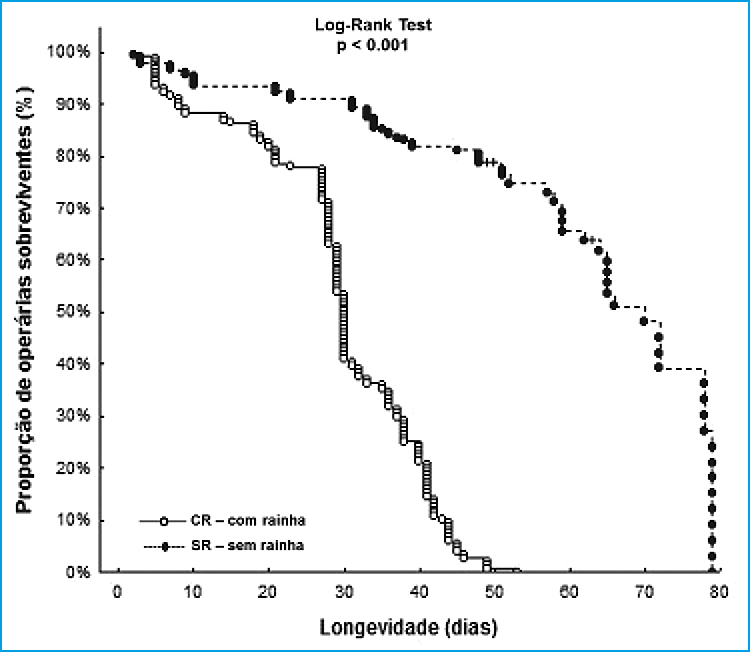
Nesse sentido, operárias de insetos sociais que evitam trabalhos arriscados podem garantir benefícios, como a oportunidade de um potencial reprodução e aumento na sua sobrevivência (Ratnieks e Reeve, 1992). Em nosso estudo verificamos que a longevidade das operárias de S. aff. postica em colônias órfãs é 33% mais longa do que das operárias que possuem rainha dentro do ninho. Trabalhos recentes (Rueppell et al., 2007; Gomes et al., 2015) mostram como o início da atividade externa influencia significativamente a longevidade das abelhas operárias, onde forrageiras precoces tem vida mais curta em relação aquelas que começam a forragear tardiamente. Em nosso estudo não foi possível observar a idade exata de início de forrageio das operárias de S. aff. postica, pois não foram feitas marcações individuais mas em coortes etárias; em colônias sem rainha as operárias em idade esperada de forrageio foram frequentemente observadas dentro do ninho, sugerindo que elas não saem todos os dias para o campo, reduzindo assim suas chances de morte e prolongando sua longevidade, esta estratégia também é sugerida por Gomes et al. (2015) para outra espécie amazônica, Melipona fasciculata.
Outra explicação para a diferença na longevidade entre operárias de colônias órfãs e com rainha é a vitelogenina (proteína precursora da gema de ovo), mas que não fazia parte do objetivo deste estudo. Em A. mellifera, foi encontrado que a retenção de vitelogenina tem influência positiva na sobrevivência das abelhas operárias, inibindo a transição comportamental para o estágio de maior risco, o forrageamento (Amdam et al., 2004; Münch and Amdam, 2010). Em abelhas sem ferrão, a maioria dos estudos focou na capacidade de oviposição de muitas espécies de meliponíneos (Dallacqua et al., 2007; Hartfelder et al., 2006). No entanto, não existem estudos em abelhas sem ferrão que tratassem do efeito da orfandade na vitelogenina e seus possíveis efeitos na longevidade das operárias.

Considerações finais
Este estudo demonstra que operárias órfãs de S. aff. postica alteram substancialmente seu comportamento, causando um aumento na sua longevidade geral e começando a colocar ovos haplóides, permitindo alguma reprodução direta pelas operárias na ausência de uma rainha.
Referências bibliográficas
Amdam, G.V., Simões, Z.L.P., Hagena, A., Norberg, K., Schrøder, K., Mikkelsen, Ø., Kirkwood, T.B.L., Omholt, S.W. (2004) Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Experimental Gerontology, 39, 767-773.
Alves, D.A., Imperatriz-Fonseca, V.L., Francoy, T.M., Santos-Filho, P.S., Nogueira-Neto, P., Billen, J., Wenseleers, T. (2009) The queen is dead – long live the workers: intraspecific parasitism by workers in the stingless bee Melipona scutellaris. Molecular Ecology, 18, 4102-4111.
Bego, L.R. (1982) On social regulation in Nannotrigona (Scaptotrigona) postica Latreille, with special reference to male production cycles (Hymenoptera, Apidae, Meliponinae). Boletim de Zoologia da Universidade de São Paulo, 7, 181-196.
Campbell, A.J., Gomes, R.L.C., da Silva, K.C., Contrera, F.A.L. (2019) Temporal variation in homing ability of the neotropical stingless bee Scaptotrigona aff. postica (Hymenoptera: Apidae: Meliponini). Apidologie, 50(5), 720-732.
Crespi, B.J. (1994) The definition of eusociality. Behavioral Psychology, 6, 109-115.
Dallacqua, R.P., Simões, Z.L.P., Bitondi, M.M.G. (2007) Vitellogenin gene expression in stingless bee workers differing in egg-laying behavior. Insectes Sociaux, 50, 70-76.
Gomes, R.L.C., Menezes, C., Contrera, F.A.L. (2015) Worker longevity in an Amazonian Melipona (Apidae, Meliponini) species: effects of season and age at foraging onset. Apidologie, 46, 133-143.
Hartfelder, K., Makert, G.R., Judice, C.C., Pereira, G.A.G., Santana, W.C., Dallacqua, R., Bitondi, M.M.G. (2006) Physiological and genetic mechanisms underlying caste development, reproduction and division of labor in stingless bees. Apidologie, 37(2), 144-163.
Imperatriz-Fonseca, V.L., Oliveira, M.A.C. (1976) Observations on a queenless colony of Plebeia saiqui (Friese) (Hymenoptera, Apidae, Meliponinae). Boletim de Zoologia da Universidade de São Paulo, 1, 299-312.
Kleinert, A.M.P. (2005) Colony strength and queen replacement in Melipona marginata (Apidae: Meliponini). Brazilian Journal of Biology, 65(3), 469-476.
Leão, K.L., Queiroz, A.C.M., Veiga, J.C., Contrera, F.A.L., Venturieri, G.C. (2016) Colony development and management of the stingless bee Scaptotrigona aff. postica (Apidae: Meliponini) using different hive models. Sociobiology, 63(4), 1038-1045.
Moraes, B.C. de, Costa, J.M.N. da, Costa, A.C.L. da, Costa, M.H. (2005) Variação espacial e temporal da precipitação no Estado do Pará. Acta Amazonica, 35, 207–214.
Münch, D., Amdam, G.V. (2010) The curious case of aging plasticity in honey bees. FEBS Letters, 584, 2496-2503.
Nunes-Silva, P., Hilário, S.D., Filho, P.S.S., Imperatriz-Fonseca, V.L. (2010) Foraging activity in Plebeia remota, a stingless bee species, is influenced by the reproductive state of a colony. Psyche, 2010, 241204.
Nunes, T.M., Oldroyd, B.P., Elias, L.G., Mateus, S., Turatti, I.C., Lopes, N.P. (2017) Evolution of queen cuticular hydrocarbons and worker reproduction in stingless bees. Nature Ecology and Evolution, 1, 0185.
Page Jr., R.E., Peng, C.Y.S. (2001) Aging and development in social insects with emphasis on the honey bee, Apis mellifera L. Experimental Gerontology, 36, 695-711.
Paxton, R.J., Bego, L.R., Shah, M.M., Mateus, S. (2003) Low mating frequency of queens in the stingless bee Scaptotrigona postica and worker maternity of males. Behavioral Ecology and Sociobiology, 53, 174-181.
Pinho, O.C., Manente-Balestieri, F.C.L., Balestieri, J.B.P. (2010) Respostas de colônias de Plebeia catamarcensis Holmberg (Hymenoptera, Apidae, Meliponina) à orfandade. Revista Brasileira Biociências, 8, 201-207.
Ratnieks, F.L.W., Reeve, H.K. (1992) Conflict in single-queen Hymenopteran Societies: the structure of conflict and processes that reduce conflict in advanced eusocial species. Journal of Theoretical Biology, 158, 33-65.
Rueppell, O., Bachelier, C., Fondrk, M.K., Page Jr., R.E. (2007) Regulation of life history determines lifespan of worker honey bees (Apis mellifera L.). Experimental Gerontology, 42, 1020-1032.
Sakagami, S.F. (1982). Stingless bees. In: Hermann HR (ed.). Social Insects. New York: Academic Press, pp 316-423.
Tan, K., Wang, Y., Dong, S., Liu, X., Zhuang, D., Chen, W., Oldroyd, B.P. (2015) Associations between reproduction and work in workers of the Asian hive bee Apis cerana. Journal of Insect Physiology, 82, 33-37.
Tóth, E., Strassmann, J.E., Nogueira-Neto, P., Imperatriz-Fonseca, V.L., Queller, D.C. (2002) Male production in stingless bees: variable outcomes of queen-worker conflict. Molecular Ecology, 11, 2661-2667.
Tóth, E., Queller, D.C., Dollin, A., Strassmann, J.E. (2004) Conflict over male parentage in stingless bees. Insectes Sociaux, 51, 1-11.
Veiga, J.C., Menezes, C., Contrera, F.A.L. (2017) Insights into the role of age and social interactions on the sexual attractiveness of queens in an eusocial bee, Melipona flavolineata (Apidae, Meliponini). The Science of Nature, 104, 31.